2025年4月22日,中國科學院遺傳與發育生物學研究所肖軍研究組在Developmental Cell在線發表了題為Dynamic control of H2A.Zub and H3K27me3 by ambient temperature during cell fate determination in Arabidopsis的研究論文,本研究綜合運用ChIP-seq、CUT&Tag、RNA-seq等技術,揭示了低溫環境通過抑制TOE1的表達及其對INO80-C復合物的招募來減緩H2A.Z組蛋白變體從染色質上的移除、同時增強PRC1復合物介導的H2A.Z單泛素化修飾。這種溫度響應的調控機制并與PRC2-介導的H3K27me3協同沉默胚胎基因,從而調控擬南芥的細胞命運。

環境溫度對植物發育至關重要,其通過激活復雜調控網絡使植物適應溫度變化。然而,不同溫度條件下染色質修飾如何精確協調細胞發育命運仍不清楚。本研究通過整合轉錄組、表觀基因組和遺傳學分析發現,在擬南芥中,較低環境溫度(16℃)可通過在異位表達的胚胎發育基因(如ABA不敏感基因ABI3和子葉發育基因LEC1)上特異性沉積H2A.Zub組蛋白修飾,部分恢復PRC2突變體中H3K27me3缺失導致的發育缺陷。這種修飾導致靶基因表達下調,從而補償H3K27me3的缺失。
研究表明:PRC1復合物催化的H2A.Zub與PRC2復合物催化的H3K27me3協同抑制胚胎基因轉錄,調控植物萌發后發育進程。低溫通過降低TOE1蛋白水平,延緩特定基因組位點H2A.Z的更新速率,從而維持胚胎基因的抑制狀態,減輕萌發后發育階段對PRC2-H3K27me3表觀遺傳標記的依賴性。該研究揭示了植物適應環境溫度變化的表觀遺傳協同調控機制,為理解環境信號與表觀遺傳網絡的互作提供了新見解。
???技術路線???
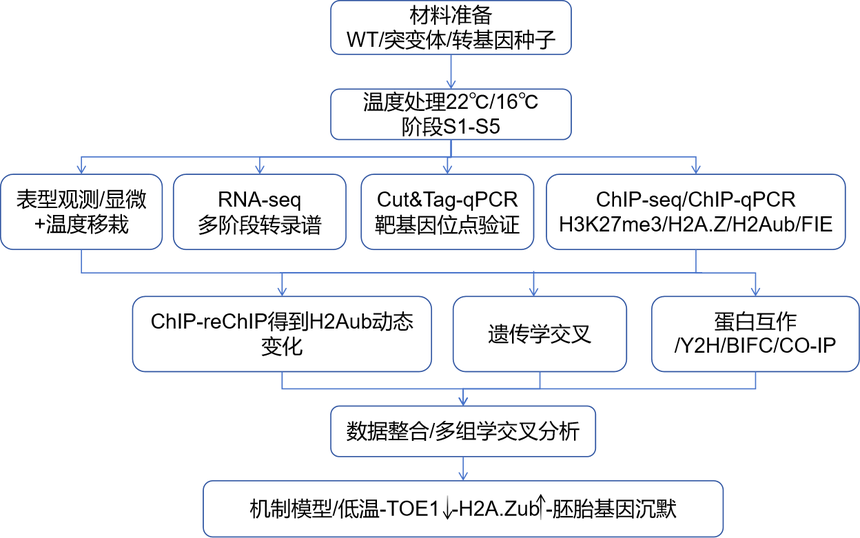
???研究結果???
1. 低溫環境可部分修復prc2突變體的發育缺陷
PRC2功能缺失(clf-28 swn-736)的擬南芥在22°C多數形成多胚葉“多克隆”組織;將幼苗降至16°C,可使≈90%以上恢復為小型但形態正常的植株(圖1A、1B)。RNA-seq比較“多克隆”與恢復植株,得到3933個差異表達基因(圖1C):胚胎/種子成熟基因(ABI3、LEC1、CUC1、LEC2)在“多克隆”中上調(圖1D、1E),光合作用-葉綠素基因在恢復植株中上調(圖1D、1F)。qRT-PCR證實,越早從22°C轉至16°C,ABI3、CUC1、LEC2表達越低,對應正常植株比例越高。ChIP-seq顯示突變體在兩溫度下H3K27me3均近乎缺失,說明低溫緩解表型不依賴該修飾。
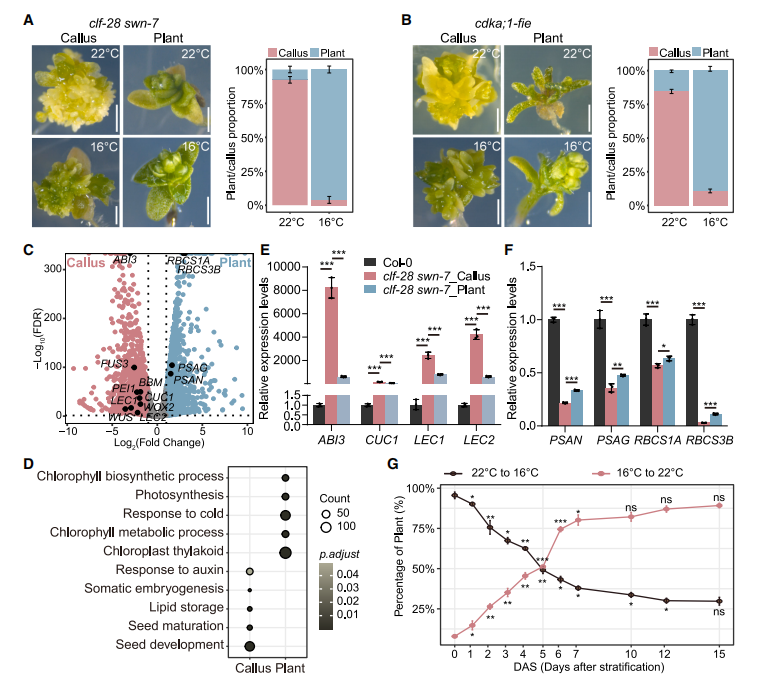
圖1:低環境溫度可部分緩解prc2突變體的發育缺陷
2.?低環境溫度抑制clf-28 swn-7中異位表達的胚胎基因
對clf-28 swn-7全發芽期分S1–S5觀察發現,表型分化始于真葉期S3,S4,對應15 DAS(從層積結束算起的天數)內細胞命運已確定(圖2A)。RNA-seq顯示S3–S5的DEGs(差異基因)數在16°C與22°C間急劇增加(圖2B),PCA亦在S3起出現溫度依賴的轉錄分離。16°C下下調的基因富集于種子發育、體胚發生,反之上調基因關聯細胞壁及葉綠體發育、減數分裂(圖2E)。與Col-0同期相比,clf-28 swn-7在16°C的轉錄組更接近野生型(圖S2C),一致于形態的部分恢復。特別是在S3-S4,有992個胚胎/干細胞相關基因(如ABI3、AIL5、CUC1、LEC1/2)在22°C上調而在16°C被抑制(圖2D、2F、2G),其低溫下的下調被認為阻止了多克隆形成。
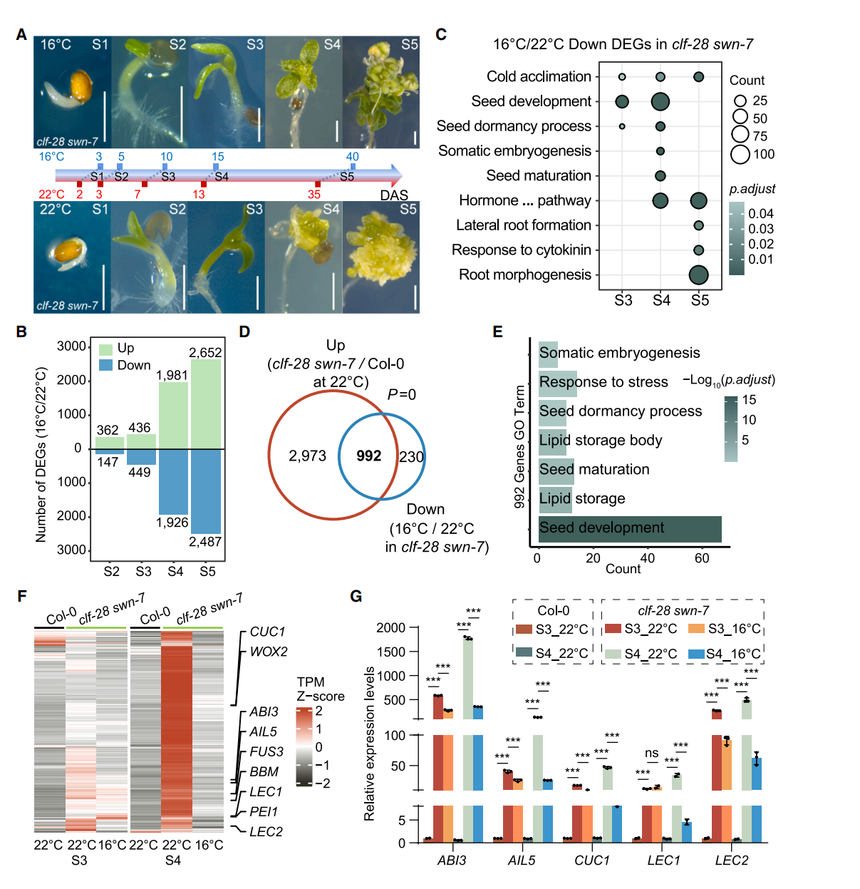
圖2. 不同溫度下clf-28 swn-7各發育階段的轉錄組分析
3.?低環境溫度降低了PRC2在靶基因上的結合
FIE(受精非依賴性胚乳)-ChIP-seq結果表明,Col-0在16°C的PRC2結合位點和強度均大幅降低(圖3A–3C),與FIE表達量無關。22°C下FIE結合的3970個基因中,僅51個(<1.3%,組I)在16°C同時上調并伴隨H3K27me3顯著下降(圖3D);其余3919基因(組II+III)雖FIE與H3K27me3均減弱,但表達保持穩定,提示存在替代性沉默機制。
clf-28 swn-7在16°C被抑制的992個基因中,341個為FIE直接靶標,并與Col-0組II+III高度重疊(222基因),富集于種子發育/體胚發生通路。代表性胚胎基因LEC1、CUC1在低溫下FIE與H3K27me3均降低卻未激活,說明低溫可借助PRC2以外的染色質修飾抑制多克隆相關基因,從而緩解突變體表型。
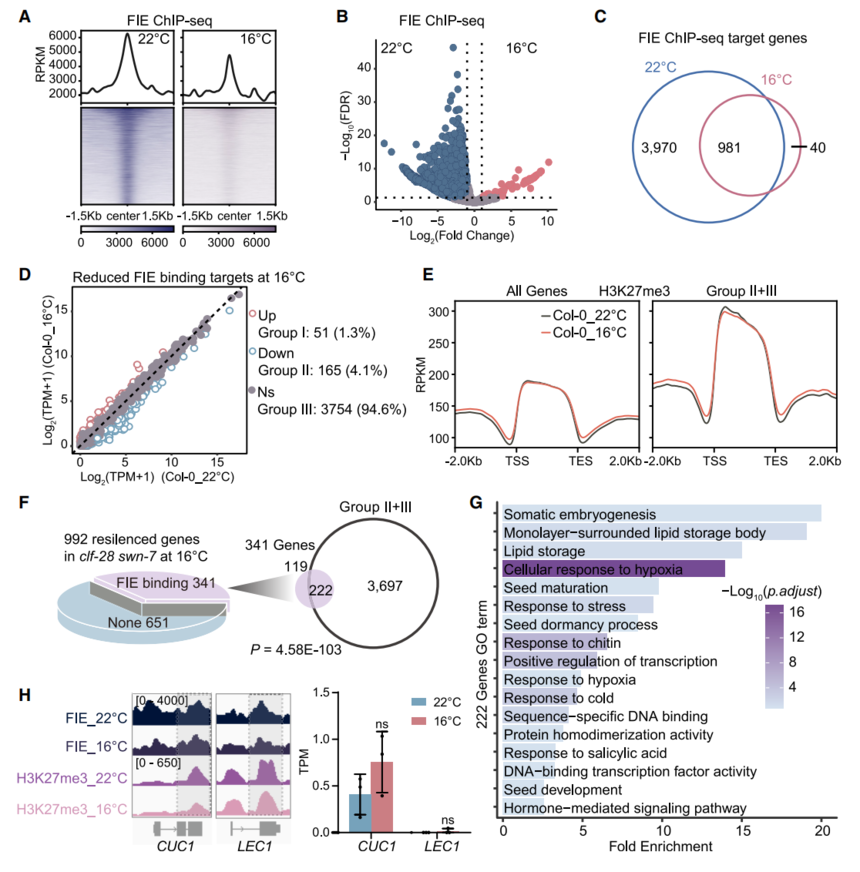
圖3. Col-0在22°C和16°C條件下FIE結合靶點的ChIP-seq分析
4.?PRC1和H2A.Z在低環境溫度下協同部分恢復clf-28 swn-7的生長缺陷
在發芽過程中,種子發育和成熟關鍵基因(ABI3、FUS3、LEC2、LEC1)依次受PRC1/PRC2抑制。為了測試PRC1是否在16°C下補償PRC2缺失,作者構建了ring1a?clf-28 swn-7三重突變體,其在16°C時僅51.3%植株表現植物形態(clf-28 swn-7為更高)(圖4A),表明PRC1必要性。ChIP-seq顯示,clf-28 swn-7中H2Aub全基因組顯著減少但在16°C略增(圖4B);而在hta10 clf-28 swn-7(H2A突變)中,16°C下植株/多克隆比例與clf-28 swn-7無異,22°C反而更高(圖4C),提示H2Aub非主要作用。相反,hta9-1?clf-28 swn-7(三重突變,H2A.Z)僅14.8%幼苗成株(圖4D)。H2A.Z ChIP-seq發現,341個基因在clf-28 swn-7中于16°C特異性富集H2A.Z(圖4E);ABI3、LEC1、CUC1的H2A.Z增加經CUT&Tag-qPCR驗證,H3K27me3未恢復(圖4F、4G)。RNA-seq與RT-qPCR顯示,這些基因在hta9-1?clf-28 swn-7中低溫下持續活躍(圖4H、4I、S4C),GO富集指向體胚發生、種子成熟/休眠。綜上,低溫下PRC1主要通過H2A.Z單泛素化而非H2Aub來強化胚胎基因沉默。
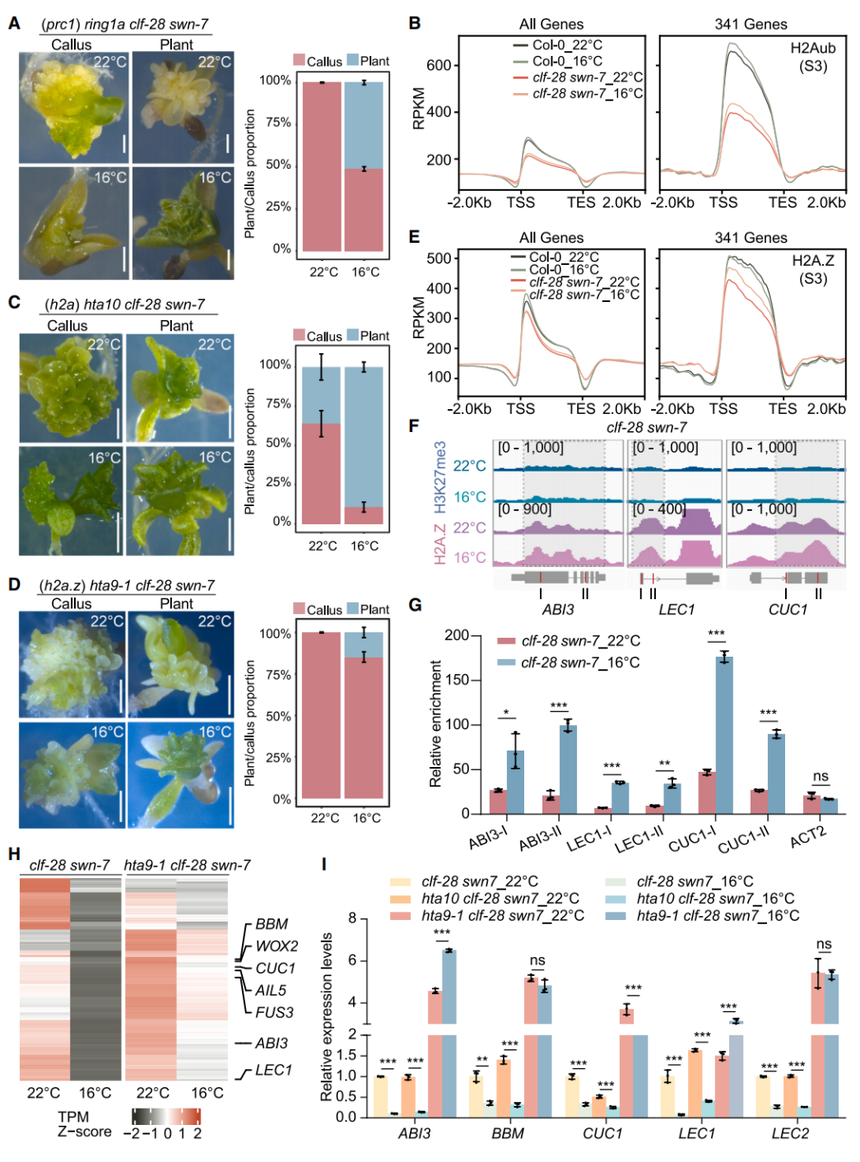
圖4. PRC1和H2A.Z(而非H2A)在低溫下部分恢復clf-28 swn-7的生長缺陷
5.?PRC1催化的H2A.Zub和PRC2催化的H3K27me3在后發芽階段協同沉默種子發育程序
hta9-1clf-28 swn-7在子葉擴展前呈現atbmi1a/b/c強突變體的白色子葉和根部膨脹表型(圖5A),其差異表達基因與atbmi1a/b/c vs Col-0 10 DAS的DEG顯著相關,提示H2A.Z與PRC1在發芽后早期協同作用。
在Col-0中,341個胚胎基因在S1→S2(2–3DAS)階段H2A.Z特異性富集后于S3(7 DAS)下降,而H3K27me3則從S1→S3逐漸增加(圖5B);ABI3、LEC1經ChIP-qPCR驗證(圖5C–5F)。
ChIP-re-ChIP顯示ABI3、LEC1、CUC1上的H2A.Zub在S1→S2增加、S2→S3減少(圖5G),其表達在h2a.z突變體S1顯著升高、在clf-28swn-7?S2→S3被激活(圖5H、S5D),表明H2A.Zub與H3K27me3協同維持發芽后胚胎基因沉默。
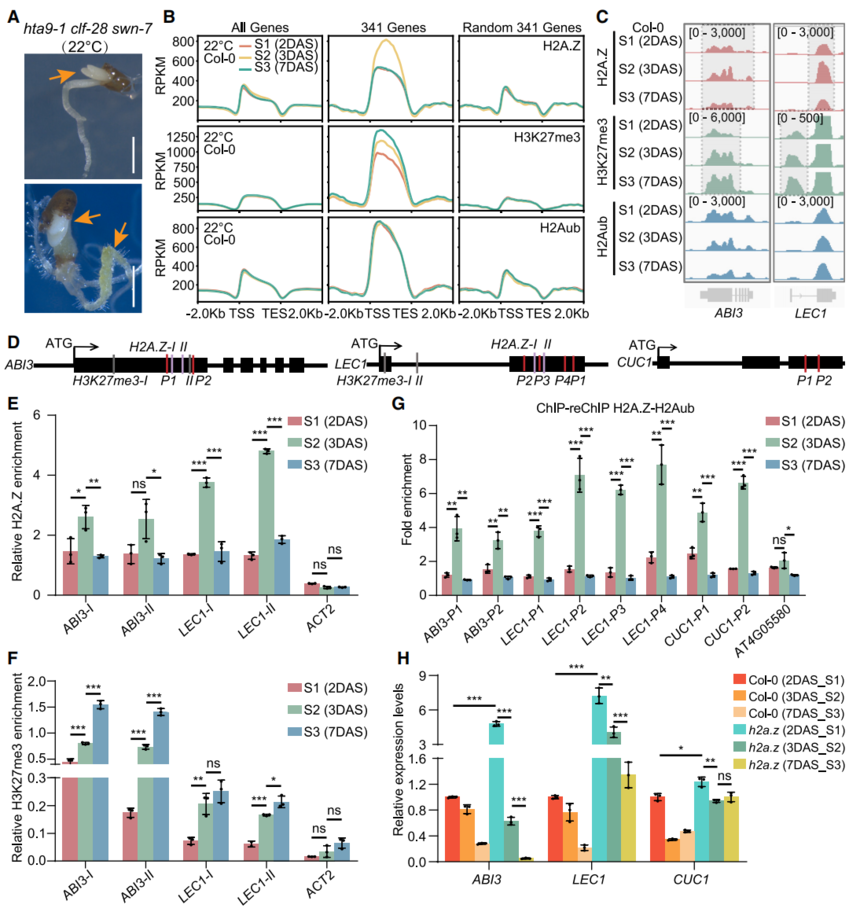
圖5:PRC1催化的H2A.Z單泛素化與PRC2催化的H3K27三甲基化協同抑制萌發后種子發育程序
6.?低溫通過 TOE1 特異性減緩 H2A.Z 的周轉速率
為探究為何低溫下clf-28 swn-7在341個細胞命運相關基因上特異性富集H2A.Z(圖4E),我們在22°C和16°C的S2、S2.5(22°C下5DAS、16°C下6DAS)及S3采樣做ChIP-seq。結果表明:在22°C,H2A.Z在S2達峰后于S2.5快速下降,S3繼續減少;而在16°C,H2A.Z從S2到S3保持高位(圖6A)。全基因組范圍內兩種溫度對H2A.Z占位的變化不顯著。
Col-0同期樣本也顯示:這341個基因在22°C H2A.Z水平顯著下降,16°C則無變化(圖5B、6B);H3K27me3在16°C下繼續累積,與22°C相似。ABI3、LEC1、CUC1實例見圖6C、6D。啟動子基序分析發現富集LEC2、TOE1、ABI3結合位點(圖6E)。Y2H、BiFC、co-IP顯示僅TOE1與INO80-C復合體成分EEN/ARP5直接互作(圖6F–6H;圖S6C、S6D)。將toe1 toe249與clf-28 swn-7交配得三重突變體,22°C下92.9%呈“半多克隆”表型(圖6I、6J)。RNA-seq和RT-qPCR顯示BBM、CUC1、LEC1、LEC2顯著下調(圖6K、6M),GO富集于體胚發生和種子成熟途徑(圖6L)。16°C下80.1%發育成植株,胚胎基因表達極低(圖6I、6M)。
以上結果表明,低溫通過TOE1調控INO80-C介導的H2A.Z周轉,從而影響關鍵胚胎基因的轉錄,部分恢復clf-28 swn-7的發育缺陷。
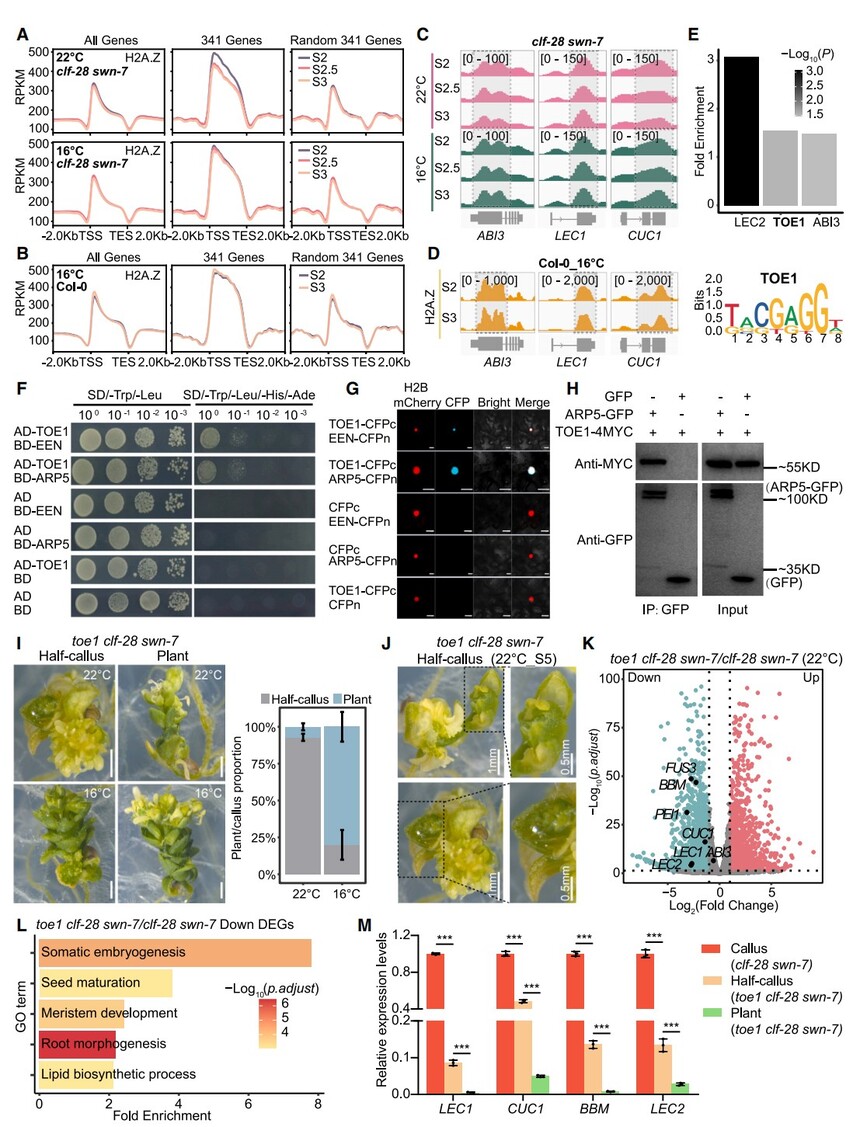
圖6. 低溫通過TOE1控制H2A.Z動態并塑造細胞發育命
7. TOE1響應環境溫度并控制胚胎基因H2A.Z的動態變化
通過ChIP-qPCR(mTOE1-FLAG)發現,22°C條件下在S2/S3階段TOE1直接結合341個胚胎基因中LEC1、LEC2和PEI1的染色質區域(圖7A、7B);在toe1 toe2雙突變體中,這些基因的H2A.Z水平在22°C下從S2到S3不降反升(圖7C、7D),而mTOE1-FLAG株系中LEC1/ABI3的H2A.Z周轉顯著加速(圖S7A),表明TOE1促進H2A.Z驅逐。盡管TOE1可通過與CLF相互作用招募PRC2(圖S7B、S7C),toe1 toe2中H3K27me3在S2–S3階段依舊累積(圖S7D、S7E),提示其沉積不受TOE1調控。此外,toe1 toe2的H2A.Z動態與低溫下Col-0相似(圖S7F),RT-qPCR和Western blot顯示16°C顯著降低TOE1轉錄與蛋白水平(圖7E、7F),而TOE1與INO80-C(ARP5)的互作強度不變(圖7G),但ChIP-qPCR證實16°C下TOE1在LEC1/LEC2/PEI1上的結合減少(圖7H),說明低溫通過降低TOE1水平減緩H2A.Z周轉。綜上,TOE1通過招募INO80-C驅逐與種子發育相關基因的H2A.Z,促進發芽后發育轉換;低溫誘導的TOE1表達下降則減緩該過程,部分挽救clf-28 swn-7的低溫發育缺陷(圖7I)。
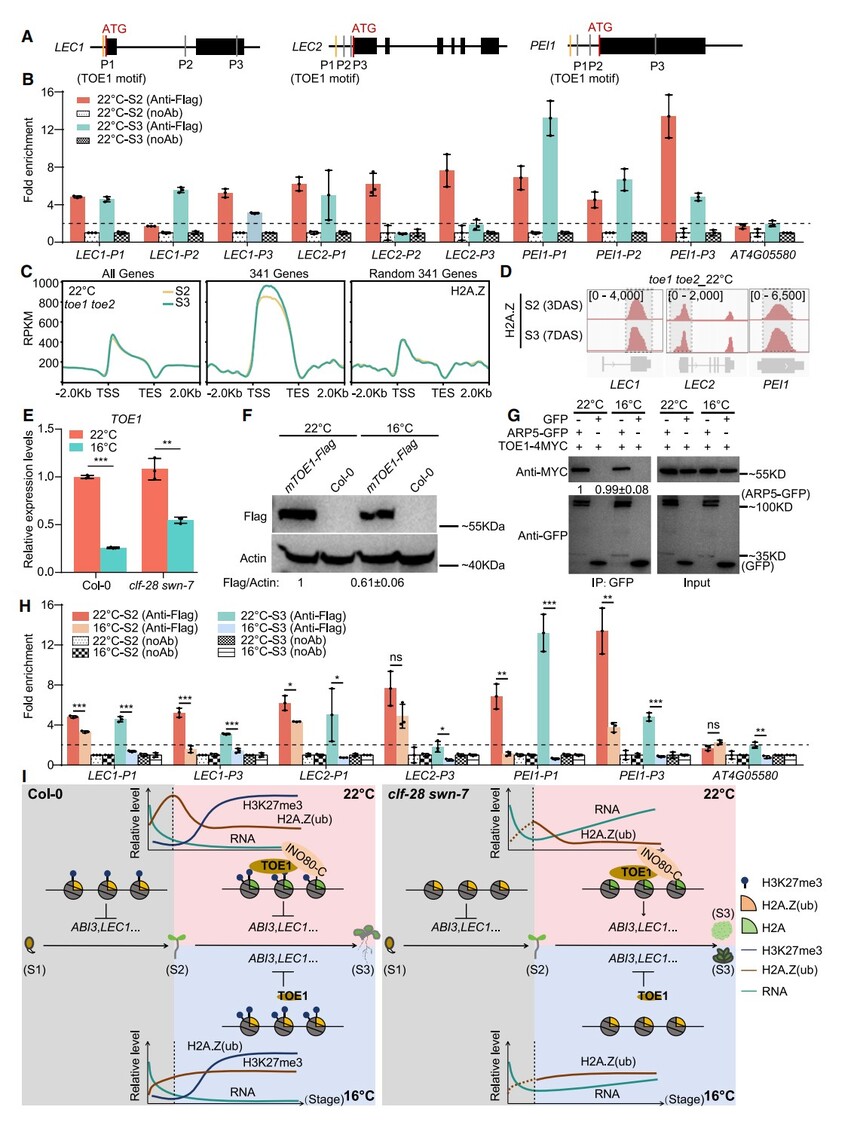
圖7. 低溫抑制TOE1表達并削弱TOE1介導的H2A.Z驅逐
該研究揭示了環境低溫(16℃)如何通過調控組蛋白變體H2A.Z的單泛素化和周轉速率,與PRC2-介導的H3K27me3協同決定擬南芥細胞發育命運。作者發現:在PRC2缺失突變體clf-28 swn-7中,低溫能促使H2A.Zub特異性積累于341個胚胎/干細胞基因,抑制其轉錄,從而部分恢復突變體的發育缺陷;這一沉積依賴PRC1而非H2A普通泛素化。正常萌發過程中,PRC1-催化的H2A.Zub先升后降,而PRC2-催化的H3K27me3逐步增加,二者接力沉默種子成熟基因。低溫通過下調轉錄因子TOE1的表達與蛋白水平,削弱其與INO80-C復合體協同驅逐H2A.Z的功能,導致上述位點H2A.Z周轉減緩并持續抑制胚胎程序;該過程與PRC2獨立。總體上,研究揭示了“TOE1-INO80-H2A.Zub”與“PRC2-H3K27me3”兩條表觀調控模塊,在溫度信號驅動下交替接管基因沉默以塑造植物的細胞命運。

)

)
![[Windows] 摸魚小工具:隱藏軟件(重制版)](http://pic.xiahunao.cn/[Windows] 摸魚小工具:隱藏軟件(重制版))





![[學習]C++ 模板探討(代碼示例)](http://pic.xiahunao.cn/[學習]C++ 模板探討(代碼示例))








