
分枝生長是作物農業特性中的一項重要指標,它直接影響植株的結構和作物的產量。黃瓜(學名:Cucumis sativus?L.)是一種在全球范圍內具有重要經濟價值和營養價值的重要蔬菜作物。在田間環境中,具有更多側枝的黃瓜植株更受青睞以提高產量。雖然光敏色素B(phyB)、分枝蛋白1(BRC1)和脫落酸(ABA)能夠促進腋芽的生長,但目前尚不清楚植物界中是否存在它們之間的整合因子。
基于此,近日中國農業大學張小蘭/趙劍宇團隊于JIPB上發表了一篇名為“The CsphyB-CsPIF4-CsBRC1 module regulates ABA biosynthesis and axillary bud outgrowth in cucumber”的文章,本文結合轉錄組、ChIP-seq、酵母雙/單雜、雙熒光素酶等實驗發現了一個涉及CsphyB-CsPIF4-CsBRC1的調控網絡,該網絡整合了光信號和ABA生物合成來調節側芽生長。這為作物育種中對分支數量的調控提供了一種策略,從而能夠實現理想的分支特性,進而最大限度地提高產量。愛基百客為本研究提供了ChIP-seq的技術服務。
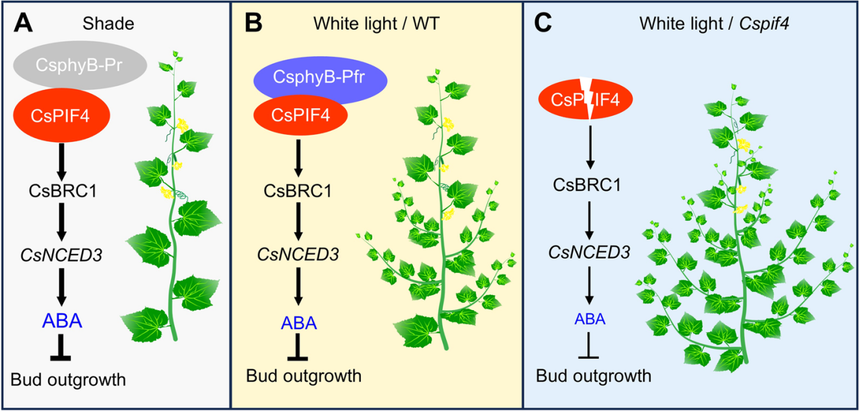
圖1CsphyB-CsPIF4-CsBRC1調節ABA合成與黃瓜側芽伸長的工作模型

? ?結果展示 ??
一.?CsphyB能夠促進黃瓜腋芽的生長
在之前的研究中,近等基因系(NIL)-lh1(其在CsphyB基因中存在一個7個堿基的缺失,導致生成的蛋白質僅含有79個氨基酸殘基)在幼苗階段與野生型(WT)Gy14相比表現出額外長的下胚軸。對成年植株的表型特征分析顯示,CsphyB突變體(NIL-lh1)的側枝明顯更少且更短,但腋芽的啟動未受影響。Gy14的腋芽逐漸伸長以形成分支,但NIL-lh1植株每個節點的枝條長度大幅縮短,并且沒有可處理的側枝(基于農業標準為長度超過4厘米的側枝)。此外,對野生型PS76進行CsphyB突變呈現出與NIL-lh1一致的表型,確定了CsphyB的功能破壞極大地阻礙了黃瓜腋芽的生長。
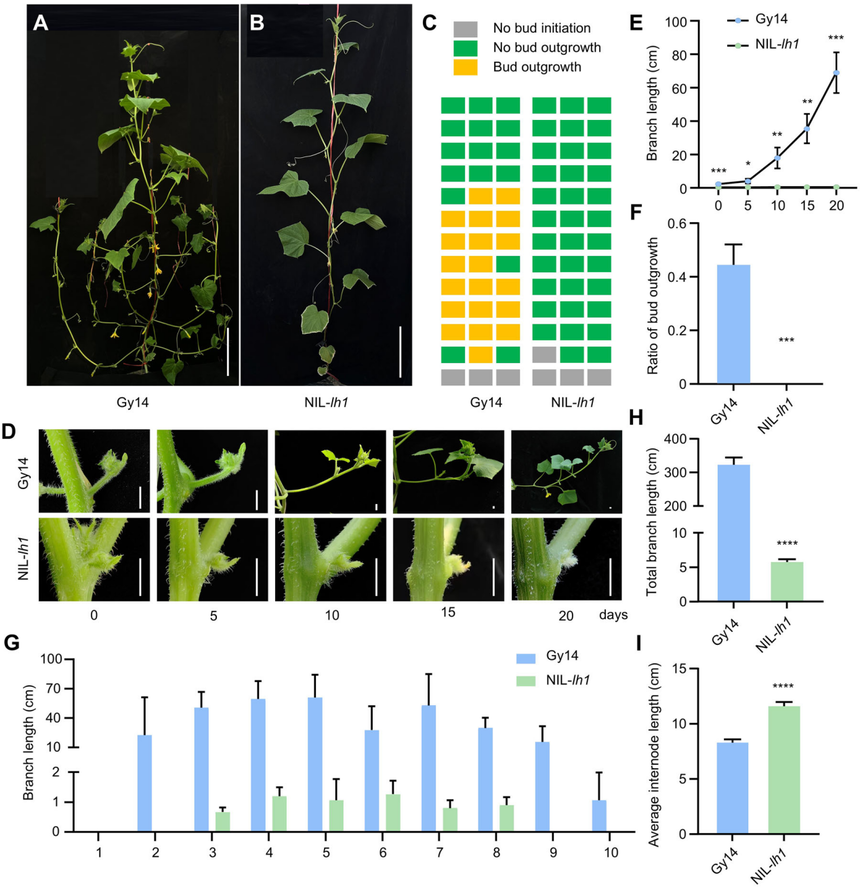
圖2Gy14(WT)和近等基因系(NIL)-lh1的表型特征分析
二. 遮光處理會抑制黃瓜的側芽生長
遮陰處理會導致遮陰逃避綜合征,表現為胚軸和側枝的伸長增加、側枝分支減少以及葉片面積減小。為了探究光信號如何調節黃瓜的側枝發育,作者對側枝過多的Sikkim inbred line WI7120進行了遮陰處理。遮陰極大地抑制了WI7120中花芽的生長,并且增加了節間長度,表明遮光抑制了黃瓜的側芽生長。
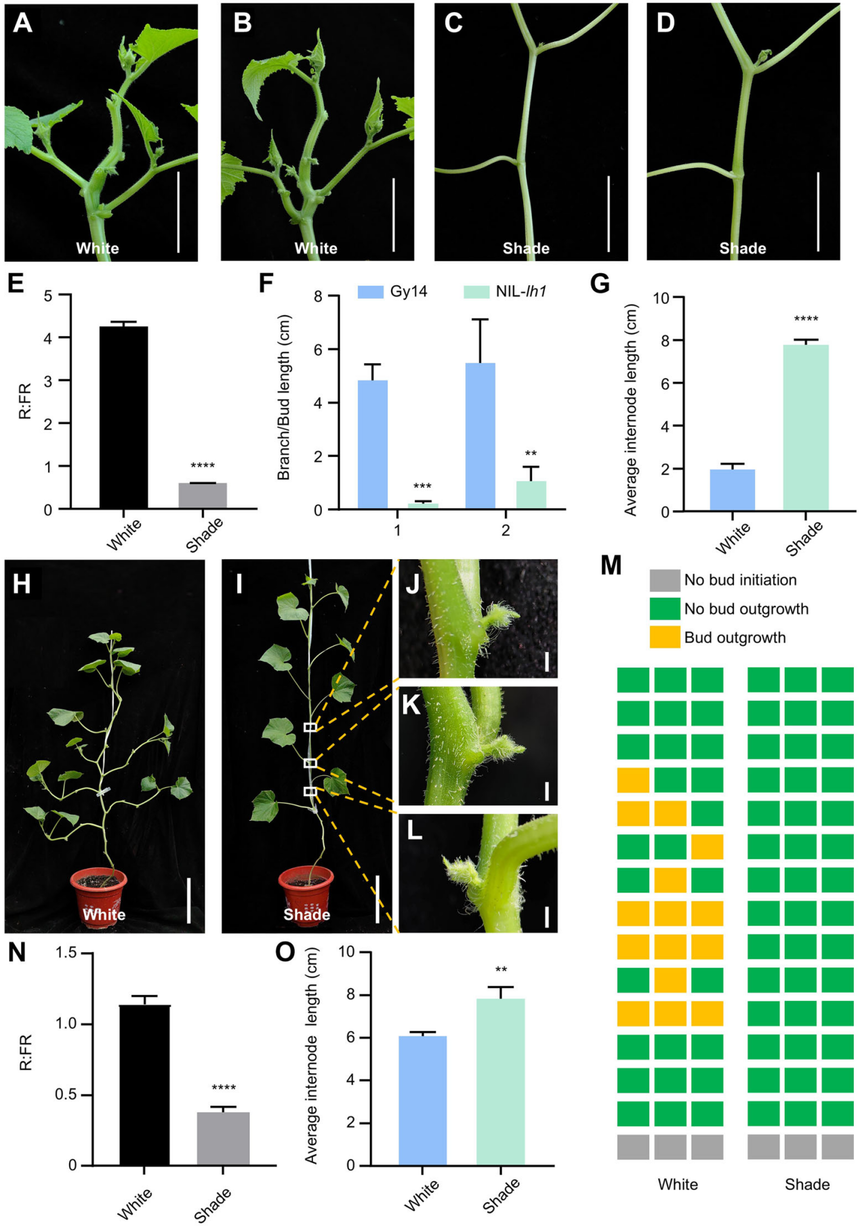
圖3遮光處理會阻礙WI7120側枝的生長
三. 脫落酸的生物合成途徑與黃瓜中由CsphyB介導的莖枝分枝現象有關
為了確定黃瓜中CsphyB的下游靶點及其調控網絡,作者使用來自Gy14和NIL-lh1同一節點的側芽進行了轉錄組測序分析。DEG分析與RT-qPCR結果顯示CsBRC1(CsGy1G003520,一種抑制黃瓜側枝生長的關鍵抑制因子),CsNCED3(CsGy4G007460)、CsNCED5(CsGy6G035690)和CsNCED6(CsGy2G009810)(三個ABA生物合成基因)在NIL-lh1中顯著上調表達。由于NCEDs在ABA生物合成的限速步驟中起作用,作者測量了Gy14和NIL-lh1同一節點側芽中的ABA含量,發現Gy14中ABA的平均濃度低于NIL-lh1,表明ABA的生物合成過程參與了黃瓜植株中由CsphyB誘導的分枝現象。
在這三種差異表達的CsNCED蛋白中,只有CsNCED3在Gy14的側芽中表達水平較高。因此,作者重點研究了CsNCED3。通過分析CsNCED3的啟動子,作者發現了一個可能的順式作用元件作為CsBRC1的結合位點。為了探究CsBRC1是否能直接調控黃瓜中CsNCED3的表達,作者使用雙熒光素酶報告系統、EMSA體外實驗進行了轉錄激活實驗驗證。為了進一步驗證CsBRC1在體內的與CsNCED3啟動子的結合,作者生成了表達35S:CsBRC1-Flag構建體的轉基因黃瓜系。染色質免疫沉淀測序(ChIP-Seq)分析顯示,在CsNCED3啟動子區域存在兩個CsBRC1結合峰,這些峰涵蓋了P1轉座子元件。這些結果有力地證明了在黃瓜體內,CsBRC1直接與CsNCED3啟動子結合。這些發現表明CsphyB可能通過CsBRC1介導的ABA生物合成來調控黃瓜的側枝生長。
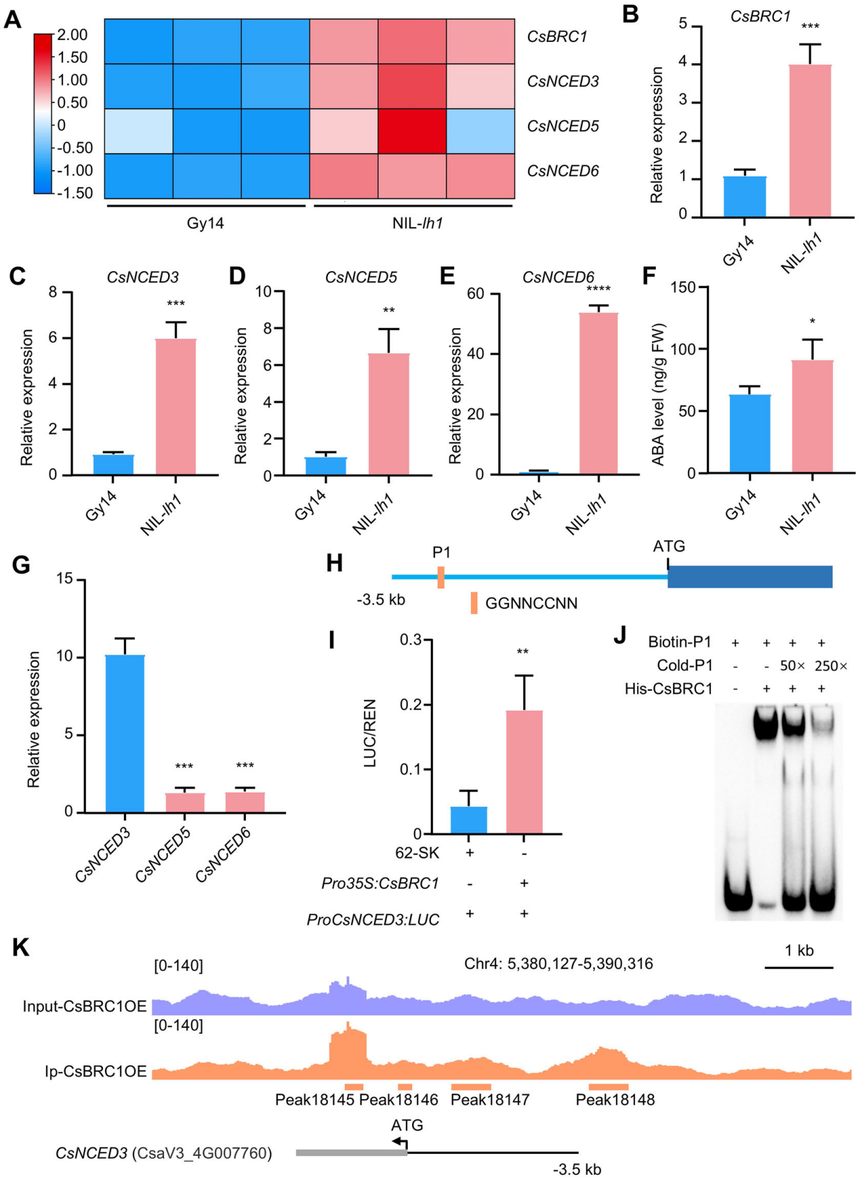
圖4黃瓜中CsphyB可能通過ABA途徑來調控黃瓜的側芽生長
四. CsPIF4與CsphyB相互作用,并直接促進CsBRC1的表達
在擬南芥中,phyB在吸收紅光后會從無活性形式轉變為有活性形式,并進入細胞核。前期研究中,黃瓜CsphyB在紅光下定位在細胞核內,并與CsPIFs相互作用。為了確認黃瓜中CsphyB與CsPIFs之間的相互作用,作者通過同源性比對搜索鑒定出了七個CsPIFs。酵母雙雜交(Y2H)實驗表明,CsPIF1a、CsPIF1b、CsPIF3和CsPIF4能夠與CsphyB相互作用,而分裂熒光素酶互補實驗驗證了CsphyB與CsPIF3/4在蛋白質水平上的相互作用。
為了探究CsPIFs是否能夠直接與CsBRC1結合,作者分析了CsBRC1起始密碼子上游的2000個堿基對的啟動子區域,并識別出了七個E-box元件,其中包括兩個典型的G-box元件(CACGTG)。酵母單雜、EMSA、雙熒光素酶實驗結果顯示只有CsPIF4能夠與CsBRC1的P1和P6轉錄激活元件結合。為了驗證CsPIF4與CsBRC1啟動子在體內的相互作用,作者生成了一個含有35S:CsPIF4-Flag構建體的轉基因黃瓜植株,并進行了ChIP實驗。染色質免疫沉淀測序(ChIP-seq)的結果顯示,CsPIF4在CsBRC1的啟動子區域存在一個結合峰,該啟動子區域包含了P6順式元件。這些結果表明,CsPIF4能夠直接與CsBRC1的啟動子結合并增強其表達。
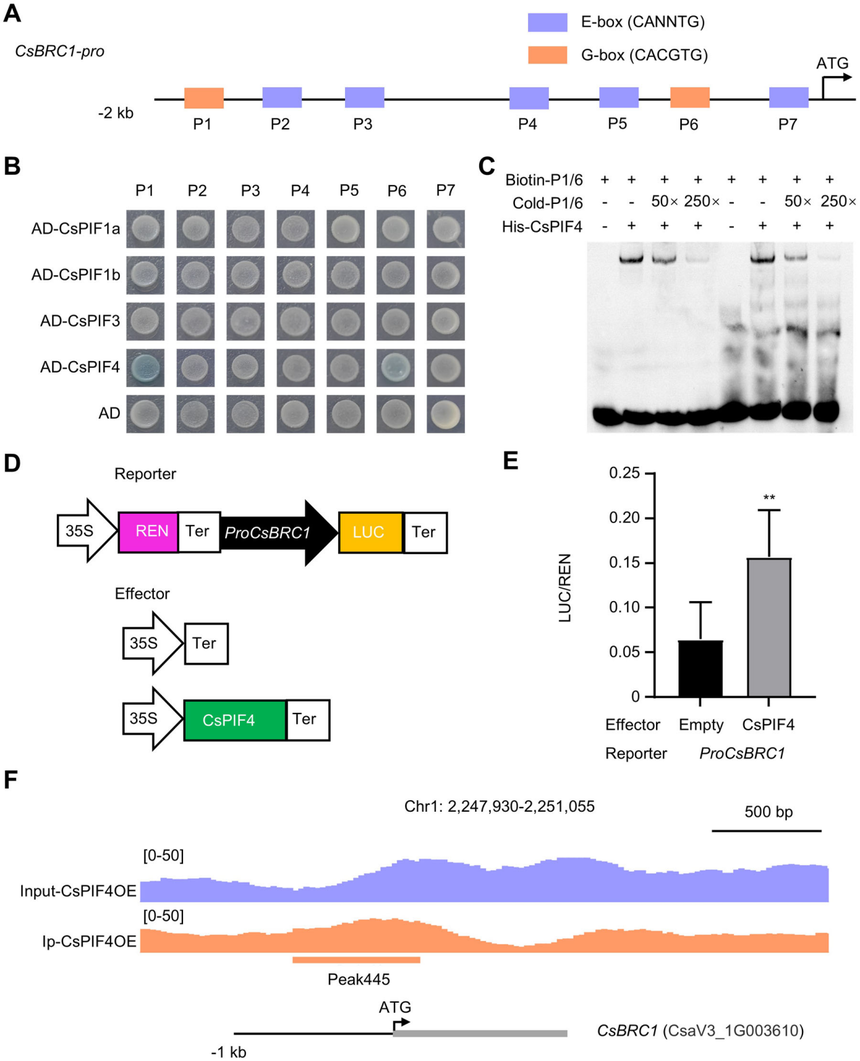
圖5CsPIF4直接與CsBRC1結合,從而激活其表達
五.?CsPIF4的功能障礙導致黃瓜側枝生長增多
為了探究CsPIF4在黃瓜中的生物學功能,作者利用CRISPR/Cas9系統生成了Cspif4突變體。獲得了兩個純合突變株,即Cspif4#1(存在84個堿基對的缺失)和Cspif4#3(存在1個堿基對的缺失),這兩個突變株分別導致蛋白質提前終止,長度分別為5個和49個氨基酸。表型特征表明,這兩個突變株的側枝數量、芽生長比例、節間分支長度均多于野生型(“9930”背景)。與野生型相比,Cspif4突變體腋芽中的CsBRC1和CsNCED3的表達水平降低,Cspif4#1突變體中的ABA濃度顯著降低。這些結果表明,CsPIF4通過刺激CsBRC1的表達和促進黃瓜中ABA的生物合成來抑制側枝的生長。
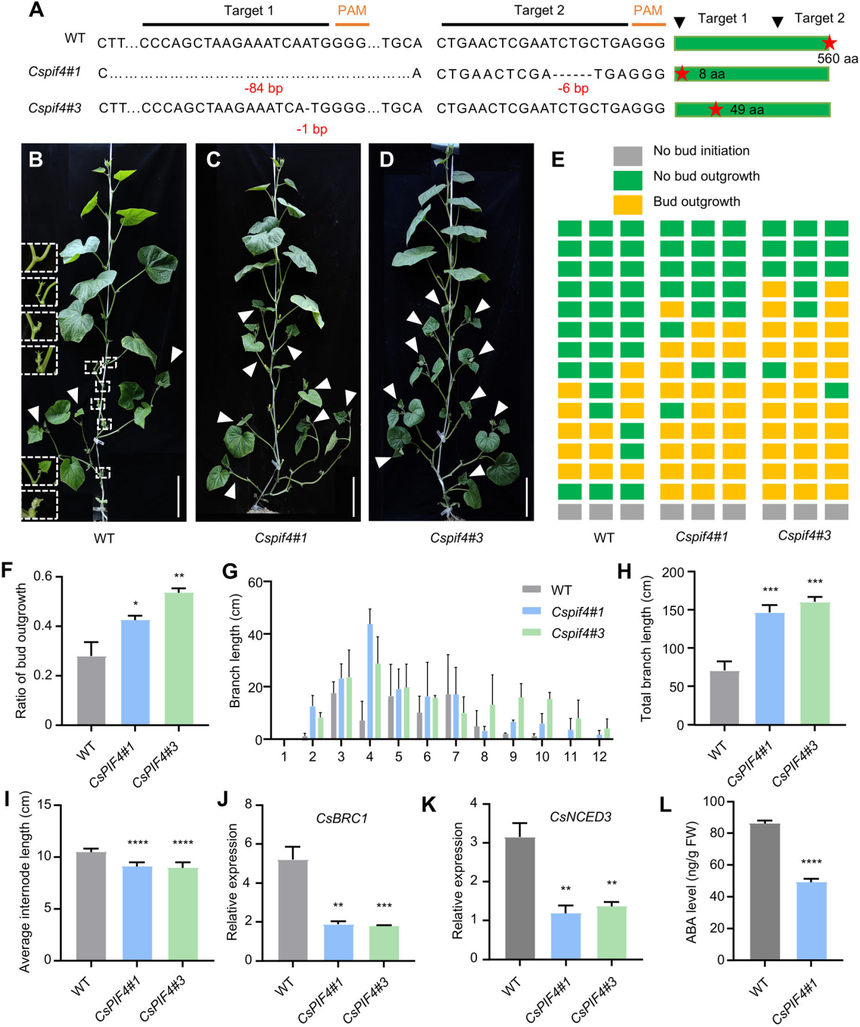
圖6黃瓜植株中CsPIF4的缺失會導致側芽的生長量增加
六.?Csnced3突變體中脫落酸的缺乏會促進側芽的生長
為了探究黃瓜中ABA積累量減少與芽生長加快之間的遺傳關系,作者利用CRISPR/Cas9技術培育了兩個Csnced3突變株系。Csnced3#1存在一個7個堿基的缺失,而Csnced3#2在第一個靶位點處存在一個1個堿基的插入;這兩種突變均導致CsNCED3蛋白翻譯的提前終止。表型分析顯示,這兩個Csnced3突變株系的側枝數量、長度,芽生長比例、均顯著大于野生型(R1461背景)。對側芽中ABA含量的進一步分析顯示,與野生型相比,Csnced3突變體中的ABA含量顯著降低。這些結果表明,Csnced3突變體中降低的ABA水平促進了側芽的生長,這表明ABA在黃瓜的側枝生長中起著負面作用。
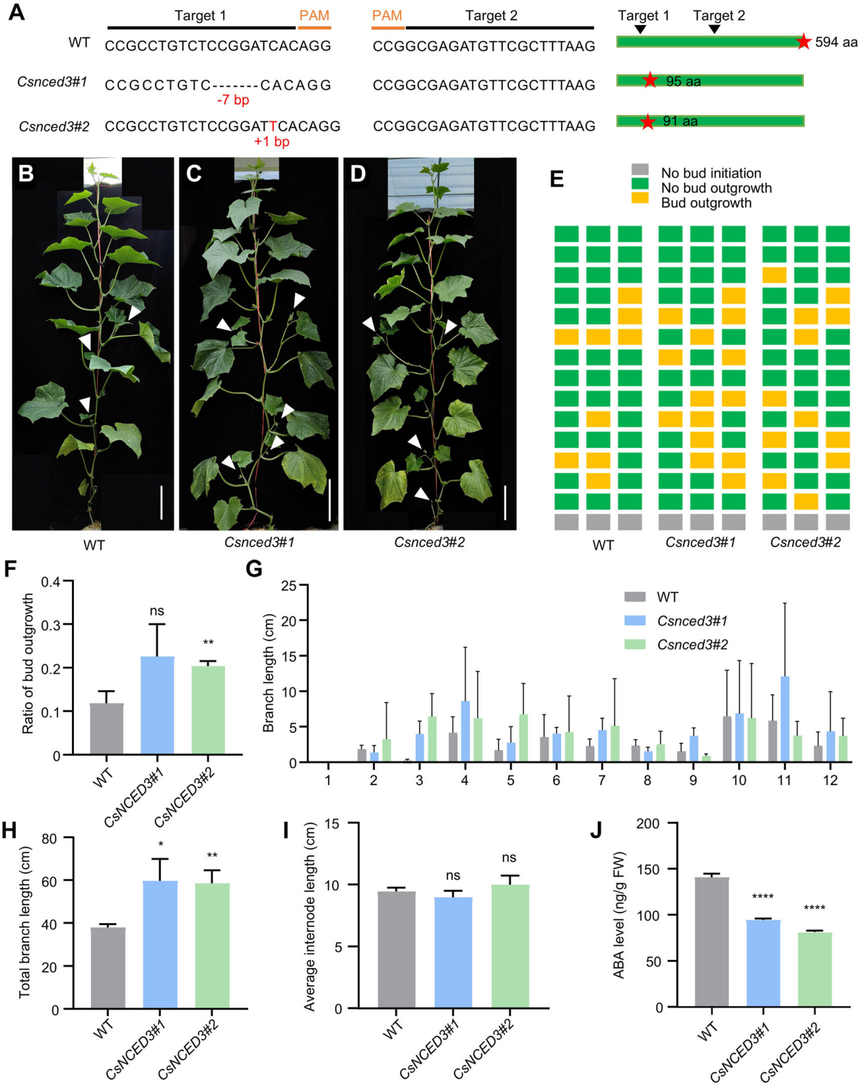
圖7黃瓜中CsNCED3的缺失導致黃瓜側芽的生長量增加
? ?總 ?結 ??
本研究以黃瓜為對象,聚焦研究了光信號與激素調控側芽萌發的分子機制。作者利用轉錄組、ChIP-seq、酵母雜交、雙熒光素酶報告基因、EMSA等實驗結合表型實驗挖掘了“CsphyB-CsPIF4-CsBRC1-ABA”的調控網絡,闡明該模塊通過整合光信號與ABA合成調控側芽萌發的機制,為作物分枝性狀改良提供育種策略。
{ 往 期 精 彩 回 顧 }
-
高分文章都在用!單細胞多組學技術和聯合分析工具介紹
-
干貨分享 ▏什么是單細胞轉錄組中的細胞通訊分析?
-
云平臺 | 玩轉單細胞比率可視化
-
小白如何挑選單細胞標記的marker基因?
精選合集,歡迎收藏喲!
-
項目文章集錦
-
ChIP專題
-
DAP-seq專題
-
ATAC-seq專題
-
單細胞測序專題
-
CUT&Tag專題
-
甲基化專題
-
RIP-seq專題
-
云平臺教程
-
國自然熱點




Linux開發工具(中)自動化構建-make/Makefile知識講解)









)
(附詳細解題思路))


![[C語言初階]結構體初階](http://pic.xiahunao.cn/[C語言初階]結構體初階)
集群技術詳解)