成熟玉米籽粒的結構
玉米籽粒的組成
成熟的玉米籽粒主要由以下三部分組成:
- 母體組織:包括種皮、胎座和花梗。種皮由珠被發育而來,起到保護種子的作用,并在種子的休眠和萌發中發揮重要作用。
- 胚:包含根分生組織、莖分生組織和5~6個小葉原基。胚根和小葉原基分別由胚根鞘和胚芽鞘包圍,起到保護作用。
- 胚乳:是玉米籽粒中最大的部分,約占總重的85%,對玉米產量具有重要意義。胚乳儲藏豐富的營養物質,為胚萌發提供營養來源。
胚乳的組織類型
成熟的玉米胚乳包含四種組織類型:
- 淀粉胚乳(Starchy endosperm):主要儲存淀粉,占玉米籽粒干物質的70%~75%。
- 糊粉層(Aleurone):在種子成熟時,是唯一包含活細胞的胚乳結構,對種子的代謝和萌發具有重要意義。
- 胚乳基部傳輸層(Basal endosperm transfer layer, BETL):在營養傳輸中起作用。
- 胚周圍區域(Embryo surrounding region, ESR):位于胚的周圍,可能在營養供給和保護中發揮作用。
玉米籽粒的營養成分
- 淀粉:占玉米籽粒干物質的70%~75%,主要儲存在胚乳中。
- 蛋白質:占10%,其中70%為貯藏蛋白,主要儲存在胚乳中。根據溶解度,胚乳蛋白可分為清蛋白、球蛋白、谷蛋白和醇溶蛋白四類。醇溶蛋白(玉米蛋白zein)占貯藏蛋白的60%以上。
- 油脂:占3.5%~6%,大部分在胚中積累。
胚乳的重要性
胚乳在成熟玉米籽粒中占據主導地位,不僅因為其在重量上的比例,還因為它儲存了大部分的營養物質,包括淀粉和蛋白質,這些對于種子的萌發和早期生長至關重要。
玉米胚的發育
早期胚胎發育的形態建成
在單子葉和雙子葉植物中,早期胚胎發育包括三個主要的形態建成過程:
- 頂-基部極性建立(Apico-basal polarity):這是胚胎發育中第一個出現的極性,決定了胚胎的頂部和底部。
- 表皮分化:表皮細胞開始分化,形成植物的保護層。
- 莖尖和根尖分生組織形成:這些分生組織是植物生長點,負責植物的進一步發育和生長。
玉米胚的發育階段
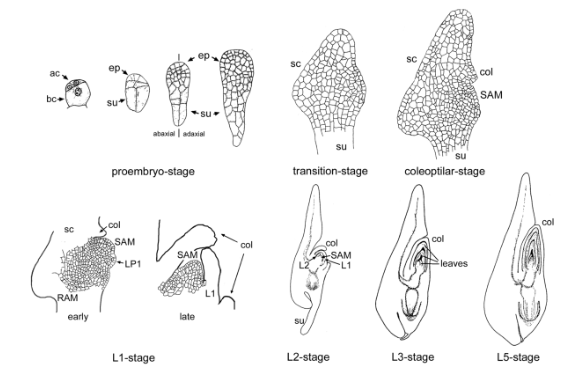
玉米胚的發育可以分為四個階段:
- 原胚期(Proembryo-stage):發生在授粉后3~5天,頂細胞和基底細胞進行有絲分裂,但維持徑向對稱發育。在原胚末期,胚形成一個幾乎沒有分化的棒狀結構。
- 轉換期(Transition-stage):在授粉后6~8天,原胚的徑向對稱發育轉變為雙側對稱性發育。此時,圍繞胚胎的一層單層外周細胞形成原表皮層,胚柄形態清晰,莖分生組織區域開始形成。
- 胚芽鞘時期(Coleoptilar-stage):在授粉后8~10天,胚芽鞘原基開始從胚近軸區突起,盾片從胚的遠軸側突起,莖尖分生組織和根尖分生組織分化形成并開始發育。
- 葉原基時期(Leaf primordium):12天后,胚芽鞘與盾片分離,在胚芽鞘下第一片葉原基形成,胚柄開始變短,成熟的胚最終形成多達五片葉原基。
玉米受精卵的第一次分裂
玉米受精卵的第一次分裂是不對稱的,分裂方向垂直于珠孔/合點軸線,形成一個小的頂端細胞和一個較大的基底細胞。基底細胞隨后轉變成一個大的囊泡細胞。
胚的近軸和遠軸面
在原胚期,胚建立近軸和遠軸面,這是胚胎發育中的重要步驟,為后續的形態建成奠定基礎。
胚的程序性細胞死亡(PCD)
在葉原基時期,胚柄發生程序性細胞死亡并開始變短,這是植物發育過程中的一種自然現象,有助于植物形態的塑造。
玉米胚乳的發育
胚乳 的早期發育包含三個主要的細胞學階段:合胞體(Coenocyte)、細胞化(Cellularization)、分 化(Differentiation)
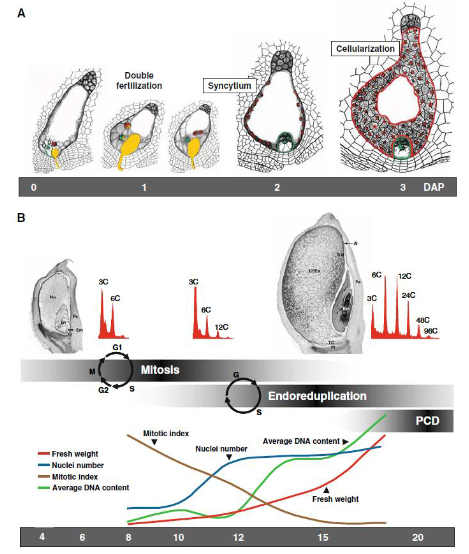
合胞體階段(Coenocyte Stage)
- 時間:受精后的72小時內。
- 過程:初生胚乳核從近胚處起始,進行連續同步的有絲分裂,產生一個由數百個游離核組成的單細胞,即合胞體。
- 特點:受精極核前三次有絲分裂產生的八個細胞核在合胞體基部圍繞胚胎的細胞質的平面上形成一個環,然后按照固定的模式向反足細胞方向遷移和重新定位,擴散到整個胚乳內側,最終產生256~512個游離核。
細胞化階段(Cellularization Stage)
-
時間:授粉后3~5天內。
-
過程:胚乳的細胞化由核表面形成的放射狀微管系統啟動,包括泡化階段和分區階段。
-
泡化階段(Alveolation Stage):
- 放射狀微管系統從每個核的核膜向外延伸,形成中間區域,為每個細胞核劃分出一個核質域。
- 細胞壁在相鄰的核質域之間沉積,核質域沿垂直于中央細胞壁的軸線方向伸長。
- 微管系統重新排列成軸向系統,一端將細胞核錨定在中央細胞壁上,另一端在細胞核頂端排列成冠狀系統。
- 成膜體在核質域邊界逐漸形成,啟動垂周細胞壁的形成,并引導垂周壁進一步向中央液泡方向生長,形成開放的管狀結構。
- 細胞壁向心伸展后,引導非姐妹核間垂周壁生長的不定成膜體和引導平周分裂的中間區域成膜體交替出現,形成次級“肺泡”。
- 泡化過程一直持續到形成2~4層細胞層。
-
分區階段(Partitioning Stage):
- 通過分割剩余的中央液泡而發生。
- 最后一個次生小泡的垂周壁持續向胚乳中心延伸,直到它們與從不同位置延伸的另一個垂周壁接觸,形成隔室。
- 垂周壁相互交匯形成的大隔室比初級小泡長,隨后它們以一種組織較差的方式進行核分裂和平周壁的生長形成較小的細胞。
分化階段(Differentiation Stage)
- 在細胞化完成后,胚乳細胞開始分化,形成不同的組織類型,如淀粉胚乳、糊粉層、胚乳基部傳輸層和胚周圍區域。
胚乳細胞分化階段
胚乳細胞分化階段可以分為兩個子階段:分化早期和分化后期。
分化早期
在分化早期,胚乳細胞主要分化出四種細胞類型:
- 胚周圍區(ESR):由具有濃密細胞質的細胞組成,細胞內液泡非常小,是胚乳中最小的細胞之一,最初在整個胚周圍明顯,后來主要在胚柄部分突出。
- 未成熟淀粉胚乳(SE):細胞較大且高度液泡化,占據胚乳的中央部分。
- 糊粉層(AL):由胚乳邊緣的第一輪泡化細胞分化而來,細胞相對較小且呈立方體形,細胞內形成多個明顯的大液泡。
- 基底胚乳傳遞層(BETL):位于胚乳的基部,毗鄰母體胎座-合點區域,細胞稍長,細胞質致密,細胞壁還未進行內增生。
分化后期
在分化后期,除了形態結構上的變化外,還分化出三種新的細胞類型:
- 亞糊粉層(Subaleurone):位于糊粉層正下方,形狀和大小與糊粉層相似,但胞質密度稍低,充滿了大的液泡、線粒體,僅有少量的蛋白體和淀粉粒。
- 傳導區(CZ):細胞非常細長,細胞核非常大,從胚乳基部向胚乳的中心部分延伸,可能起著運輸營養物質的作用,胞質密度比相鄰的BIZ和SE細胞低。
- 基底中間區(BIZ):緊鄰BETL上部的2~4層細胞,形態上明顯區分于BETL,處于BETL和CZ細胞的中間狀態,基因表達分析表明它們是三種不同細胞類型。
細胞形態和功能的變化
- ESR細胞:在分化后期變長,空泡化程度更高。
- 淀粉細胞核:大小從小細胞向積累貯藏化合物的較大的內層淀粉細胞逐漸增大。
- 糊粉層細胞:保持方形,小液泡變少,胞質致密并有脂類堆積。
- BETL細胞:明顯變長,細胞壁內增生明顯,質膜表面積增加,細胞質致密且含有大量液泡。
研究進展
- 籽粒結構和發育機制:通過對大量玉米籽粒突變體的研究,科學家們對玉米籽粒的結構和發育機制有了更深入的了解。
- 基因克隆和驗證:利用突變體,科研工作者克隆并驗證了眾多調控籽粒發育的基因,包括:
- 淀粉合成相關基因
- Pentatricopeptide repeat(PPR)蛋白編碼基因
- 玉米胚乳基底傳遞層(BETL)發育相關基因
- 生長素和信號轉導相關基因
- 蛋白體和淀粉粒發育相關基因
突變體類型及其影響
這些基因的突變會導致玉米籽粒出現不同程度的發育缺陷,主要分為以下幾種類型:
- 缺陷型籽粒(defective kernel,dek)突變體:籽粒發育不完全。
- 空果皮(empty pericarp, emp)突變體:胚和胚乳均不能發育。
- 胚特異性突變體(embryo-specific,emb):影響胚的發育。
- 小粒突變體(small kernel,smk):籽粒體積減小。
- 胚乳特異性突變體:
- 影響貯藏蛋白和淀粉生物合成的突變體(如opaque;floury)。
- 影響糊粉層分化和色素形成的突變體。
PPR蛋白的基本特征
- 結構:PPR蛋白由35個氨基酸的重復序列串聯排列構成,具有簡并性。
- 分類:PPR蛋白分為P型和PLS型兩大類。
- P型:只含有35個氨基酸的典型P基序。
- PLS型:由P、L(35或36個氨基酸)和S(31或32個氨基酸)基序組成PLS三聯體重復串聯排列。PLS型蛋白的羧基末端有高度保守的E、E+或DYW結構域,進一步分為PLS、E、E+和DYW亞類。
PPR蛋白在植物中的功能
- 數量和功能:植物中PPR蛋白成員數量龐大,通常以非冗余的方式發揮功能,參與多種生物學過程,如光合作用、葉片發育、植物生長、種子發育、脅迫反應、花粉育性等。
- 定位:大多數PPR蛋白定位于線粒體或葉綠體。
PPR蛋白在玉米籽粒發育中的作用
- 影響:PPR蛋白的變異可以導致胚和胚乳發育缺陷,產生不同類型的突變體,如缺陷型籽粒(dek)、空果皮(emp)、胚特異性(emb)、小粒(smk)等。
- 葉綠體定位的PPR蛋白:通常導致胚胎發育缺陷,產生emb類型突變體。
- 線粒體靶向的PPR蛋白:突變會對胚和胚乳的發育造成不同程度的影響,產生多種類型的突變表型。
PPR蛋白的具體功能
- P型PPR蛋白:主要負責線粒體內含子的剪接,如Dek2、Emp8、Emp11等參與線粒體不同基因內含子的剪接。
- PLS型PPR蛋白:主要負責線粒體內含子的編輯,且每個基因通常可以對多個位點進行編輯。
其他PPR蛋白的功能
- PPR8522、EMB-7L和qKW9:這些PPR蛋白參與葉綠體RNA的轉錄、剪接和編輯。
BETL的功能
- 營養物質運輸:BETL是母體向子代運輸營養物質的入口,在籽粒灌漿過程中起著至關重要的作用。
BETL相關基因及其作用
-
ZmMRP1 (Myb-Related Protein-1):
- 表達:在玉米BETL中特異性表達。
- 功能:作為BETL細胞特異性的轉錄激活因子,調節BETL-1、BETL-2、BETL-9等基因的表達,促進BETL細胞的分化并維持其形態建成。
-
ZmMn1:
- 編碼:編碼細胞壁轉化酶INCW2。
- 定位:INCW2蛋白錨定在BETL細胞內增生細胞壁上。
- 功能:INCW2功能損失導致BETL中游離質外體己糖缺乏,胚乳不能獲得充足的糖類,突變體粒重顯著降低。
-
ZmSWEET4c:
- 功能:通過內增生的BETL細胞向胚乳轉運葡萄糖和果糖。
- 影響:ZmSWEET4c功能缺陷導致胚乳灌漿嚴重不足,但胚能發育為正常植株。
-
ENB1:
- 編碼:編碼纖維素合酶5(ZmCESA5)。
- 表達:主要在胚乳基底傳遞層細胞中表達。
- 功能:ENB1功能受損導致BETL細胞壁內增生劇烈減少,BETL細胞向胚乳運輸糖的功能受損,阻礙胚乳中淀粉的合成,并導致胚乳中淀粉過早被發育中的胚胎利用,產生胚乳分解的籽粒表型。
基因表達調控
- 糖信號調控:ZmMn1和ZmSWEET4c的表達很可能受BETL細胞中糖信號的調控,并進一步誘導ZmMRP1的表達。
研究意義
- BETL細胞內增生:ENB1的研究提供了BETL細胞內增生促進糖分向胚乳運輸的直接證據。
- 籽粒灌漿:這些基因的功能對于理解BETL在籽粒灌漿過程中的作用至關重要,有助于揭示籽粒發育的分子機制。
總結
這段文字強調了BETL在玉米籽粒發育中的關鍵作用,以及ZmMRP1、ZmMn1、ZmSWEET4c和ENB1等基因在BETL細胞形態建成和功能中的重要性。這些基因的表達和功能對于BETL細胞的分化、細胞壁形成、糖分運輸等過程至關重要,影響籽粒的灌漿和最終的粒重。對這些基因的研究有助于我們更好地理解植物種子發育的分子機制,為作物改良和農業生產提供理論基礎。




)





——網絡問題)



:一文詳解three.js中的矩陣Matrix和向量Vector)

:常用方法、遍歷、推導式與嵌套實戰)


